

研究成果・プレスリリース
【プレスリリース】「赤と緑はなぜ見分けられるのか」霊長類色覚の分子構造を解明 ~赤・緑錐体視物質の構造を原子レベルで決定、30ナノメートルの謎に迫る~
発表のポイント
- 世界初:霊長類の「赤」と「緑」の視物質の立体構造を原子レベルで解明
- わずか3つのアミノ酸が「色の見分け」を支配する仕組みを特定
- 錐体特有の「横穴」構造を発見:迅速な視覚再生の鍵を提示
- 色覚多様性の理解から次世代の創薬・治療戦略への応用に期待
概要
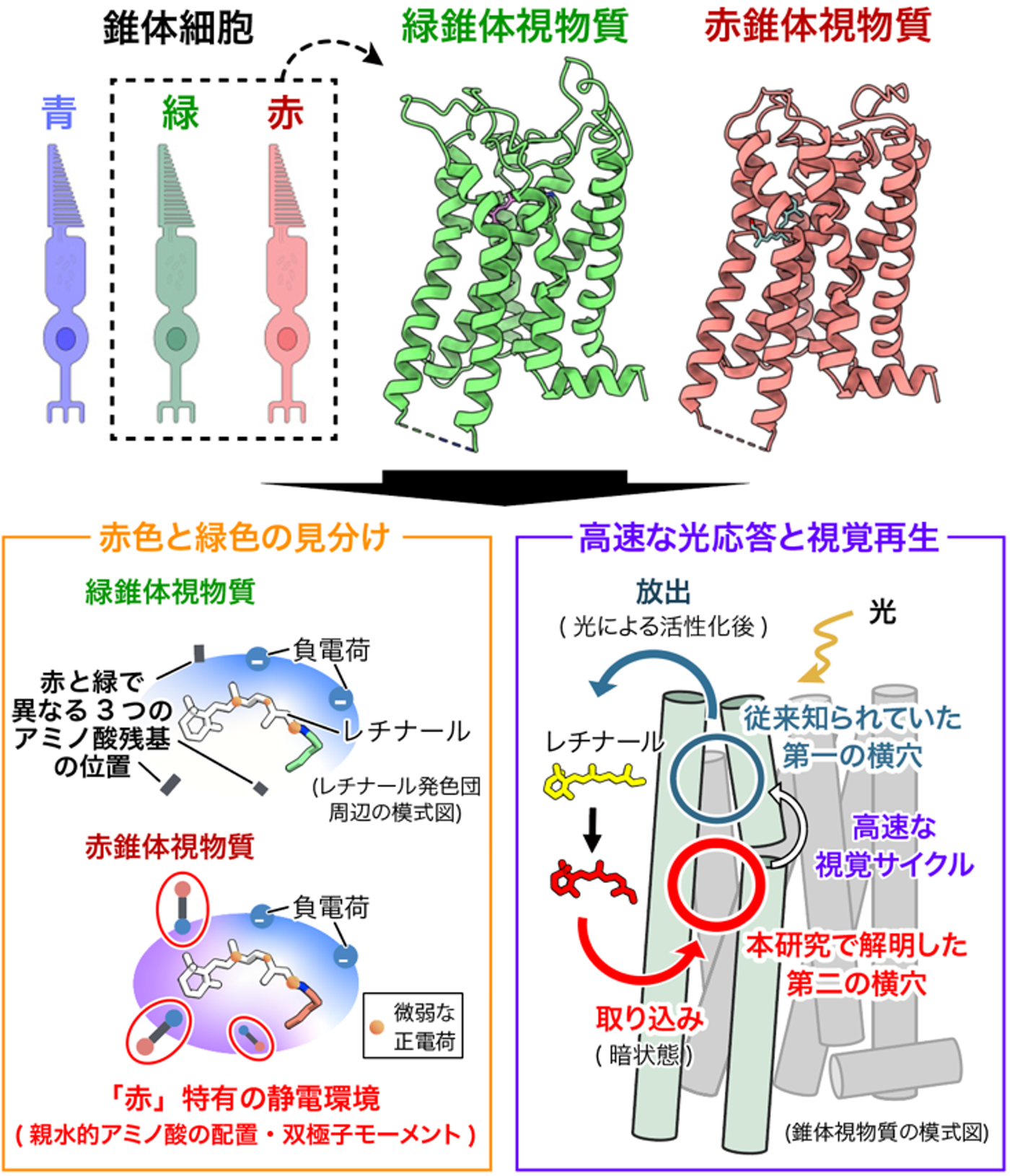
研究の背景
研究の内容・成果
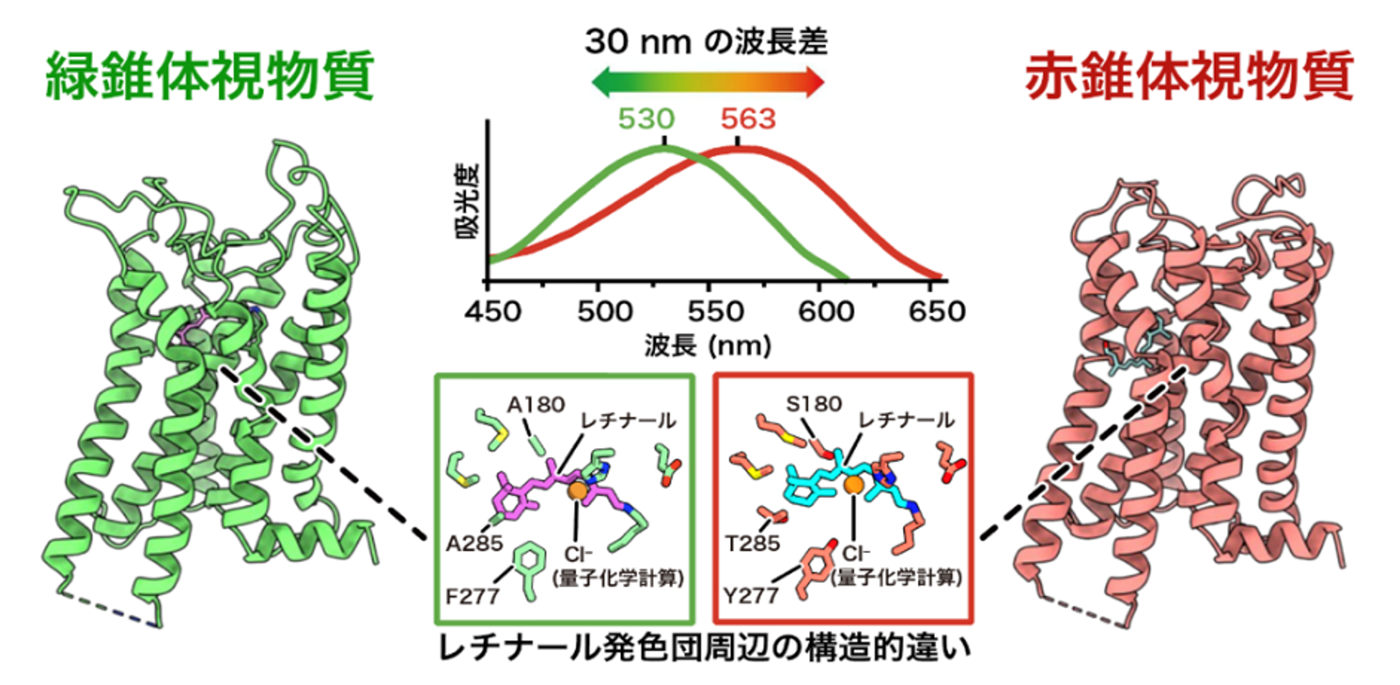
本研究グループは、最先端のクライオ電子顕微鏡単粒子解析を用い、霊長類 (マカク) の赤および緑錐体視物質の構造を、分解能3.2〜3.7 Å (オングストローム) で決定することに世界で初めて成功しました。この構造決定には、タンパク質構造予測ツール (AlphaFold2) と独自アルゴリズムを組み合わせたコンストラクト最適化プログラム (NOAH) (注7) を活用し、構造解析に適した試料設計を効率的に行ったことが大きく寄与しています。これにより、光を受ける前の「暗状態」における視物質の精密な三次元構造が明らかになり、長年謎であった色覚の分子基盤へ直接迫ることが可能となりました。得られた構造を詳細に比較した結果、光を吸収する中心分子である「レチナール」自体の化学構造は両者でほぼ同一である一方、赤錐体に特異的な「3つの親水性アミノ酸残基」がレチナール周辺に形成する構造的環境の違いが、吸収波長を長波長側 (赤側) へとシフトさせる決定的な要因であることを突き止めました。
この波長制御の仕組みをさらに深く検証するため、赤・緑視物質間でアミノ酸を入れ替えた変異体実験と、三次元構造に基づく精密な量子化学計算を統合して解析を行いました。その結果、これら3つの残基による影響は、レチナールとの単純な距離だけでは説明できず、アミノ酸側鎖の「向き」や「電気的な性質 (双極子モーメント)」といった空間的な静電環境に強く依存していることが判明しました。すなわち、原子レベルで緻密に設計されたタンパク質内部の静電的な差異こそが、赤と緑を分ける「約30 nm」という絶妙な波長差を生み出している実態を、世界で初めて構造基盤に基づいて証明しました (図1)。
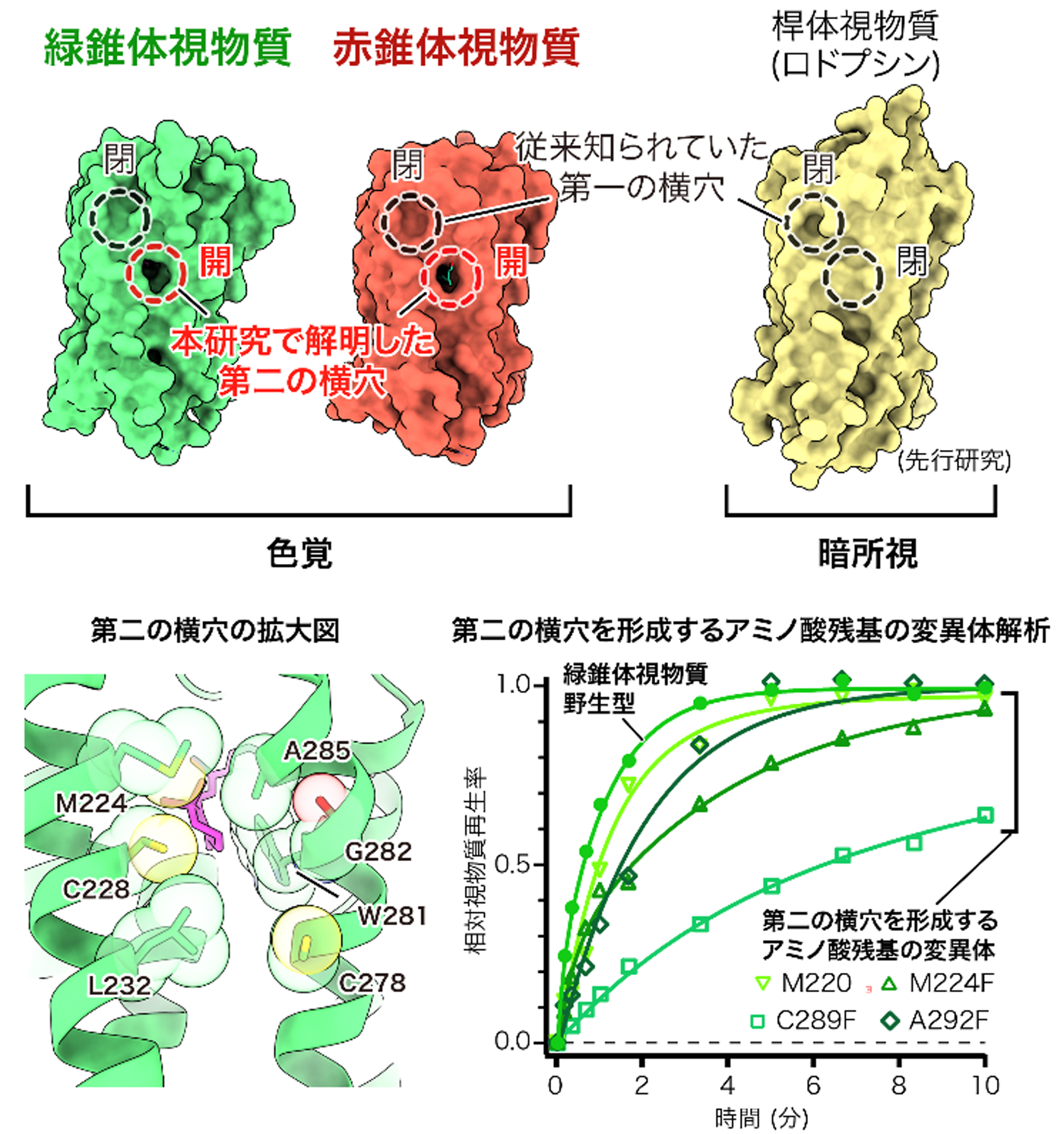
社会的な意義・今後の展望
本研究は、霊長類が持つ精緻な色覚の仕組みを、立体構造という物理的な証拠に基づいて初めて明らかにしたものです。アミノ酸配列が約96%共通していながら、わずか数個のアミノ酸の配置や電気的な性質の違いが、赤と緑を分ける「30 nm」の波長差を生み出す。この洗練された分子メカニズムを原子レベルで統合的に説明した点に、生命科学における極めて深い意義があります。
用語説明
ヒトを含む霊長類の網膜にある「錐体細胞」に存在し、色を感知するタンパク質。赤・緑・青の3種類があり、それぞれが特定の波長の光に反応することで、脳が色を識別します。一方、暗い場所で働く視物質は「桿体視物質 (ロドプシン)」と呼ばれます。
タンパク質などの生体分子をマイナス180℃以下の急速凍結状態で観察し、電子顕微鏡で撮影した多数の分子像から、その3次元構造を決定する手法。結晶化が困難な不安定なタンパク質でも、原子レベルの精密な構造を捉えることが可能です。
量子力学の原理に基づき、分子内の電子の振る舞いを計算することで、物質の性質を予測する手法。本研究では、タンパク質内部の複雑な静電環境が、レチナールの吸収する光のエネルギー (波長) にどのような影響を与えるかを精密に解析するために用いられました。
ビタミンAから作られる、光を受け取るための小さな分子。視物質タンパク質の内部に結合しており、光を吸収するとその形を変化させます。これがスイッチとなり、タンパク質全体の構造変化を引き起こして視覚信号が発生します。
分子やアミノ酸残基内部における、プラスとマイナスの電荷の偏りの大きさと方向を示す指標。本研究では、赤錐体特有のアミノ酸が持つこの「電気的な偏り」の向きが、レチナールとの相互作用を通じて波長を長波長側へシフトさせていることを突き止めました。
タンパク質分子の内部から外部 (膜側面) へと通じている小さな隙間。本研究では、錐体視物質において、光によって変性したレチナールを排出し、新しいレチナールを迅速に取り込むための「搬入口」として機能している可能性が示されました。
タンパク質構造予測 (AlphaFold2) を活用し、クライオ電子顕微鏡解析に適したタンパク質コンストラクトを効率的に設計するために開発された独自プログラム (NOAH:NOn-experimental, AI-assisted High-throughput construct screening)。
本手法の詳細は関連論文 (https://doi.org/10.64898/2026.04.02.716066) に報告されています。
細胞膜に存在し、ホルモンや光などの外部刺激を細胞内へと伝える役割を持つ受容体の総称。ヒトには約800種類存在し、その重要性から現在市販されている医薬品の約3割がこのGPCRを標的としています。視物質もこのファミリーの代表的な一員です。
論文情報
Structural insights into spectral tuning and retinal exchange in cone visual pigments
<著者名>
<掲載雑誌名>
<DOI>
本研究は、「JST 創発的研究支援事業 (課題番号:JPMJFR240U, JPMJFR204S)」、「JST 戦略的創造研究推進事業 CREST (課題番号:JPMJCR25B5, JPMJCR1753, JPMJCR21P3, JPMJCR23B1)」、「JST 戦略的創造研究推進事業 さきがけ (課題番号:JPMJPR19G4, JPMJPR24OF)」、「JST 戦略的創造研究推進事業 ACT-X (課題番号:JPMJAX222F)」、「JSPS 科学研究費助成事業 (課題番号:JP20H05440, JP21H04969, JP22H00400, JP25H01338, JP 24H02262, JP25K09525, JP26H00459, JP26H00461, JP19H03428, JP24K02231, JP24K18060, JP25H02243, JP21H05111, JP21H05112, JP26K18246, JP25KJ1446, JP24KJ0981)」、「AMED 再生・細胞医療・遺伝子治療実現加速化プログラム (課題番号:JP24bm1123057)」、「AMED-BINDS 生命科学・創薬研究支援基盤事業 (課題番号:JP23ama121001, JP23ama121002, JP24ama121001, JP24ama121002)」、「AMED 創薬基盤推進研究事業 (課題番号 JP26ak0101309h0001)」、「文部科学省・学際領域展開ハブ形成プログラム・マルチスケール量子-古典インターフェース研究コンソーシアム (課題番号:JPMXP1323015482)」、「京都大学ヒト行動進化研究所 共同利用研究プロジェクト」、「京都大学研究連携基盤」、「中山人間科学振興財団」、「中島記念国際交流財団」、「光化学技術研究振興財団」、「アステラス病態代謝研究会」、「風戸研究奨励会」などの支援を受けて行われました。
問い合わせ先
【広報担当】
東北大学 多元物質科学研究所 広報情報室
TEL: 022-217-5198
E-mail:press.tagen*grp.tohoku.ac.jp
お問い合わせ
東北大学 国際放射光イノベーション・スマート研究センター
〒980-8572 仙台市青葉区荒巻字青葉468−1
